| Contributors | Affiliation | Role |
|---|---|---|
| Caron, David | University of Southern California (USC) | Principal Investigator |
| Fu, Feixue | University of Southern California (USC) | Co-Principal Investigator |
| Hutchins, David A. | University of Southern California (USC) | Co-Principal Investigator |
| Schnetzer, Astrid | University of Southern California (USC) | Co-Principal Investigator |
| Tatters, Avery | University of Southern California (USC) | Contact |
| Groman, Robert C. | BCO-DMO Data Manager |
Experimental design
A flow chart of the experimental design is shown in Figure 1, including collection and incubation of the natural bloom in a short-term (2 week) pCO2 experiment, isolation of cells of all four species from each pCO2 treatment, conditioning of the isolates at the pCO2 from which they were isolated for one year, and recombining isolates into artificial communities to compete for 2-week periods following 4, 8 and 12 months of conditioning. Growth rates of each species were assessed during conditioning in unialgal cultures at the 8-month timepoint, as well as in mixed communities during the initial bloom experiment and the 12-month artificial community experiment. “Switch” competition experiments in which clones conditioned at each pCO2 were competed in artificial communities at the other two pCO2 levels were also performed after 12-months. The switch competition data are in a separate data set.
Initial pCO2 incubation experiment
A mixed natural dinoflagellate bloom dominated by Lingulodinium polyedrum, Prorocentrum micans, Alexandrium sp. and Gonyaulax sp. at a total cell density of ~700 cells per ml was collected off Venice Beach, California in September, 2009. This large regional bloom extended throughout the Southern California Bight region. Samples were collected near shore for both the initial incubations and all experimental dilution water used throughout the 12-month experiment.
The experiment was incubated in the laboratory at 18 degrees C under 90 photons per meters squared per second of cool white fluorescent illumination on a 14-h light: 10-h dark cycle. Triplicate sterilized 1 L polycarbonate bottles were gently bubbled (60 bubbles per minute) using commercially prepared air/CO2 mixtures (230 ppm, 433 ppm and 765 ppm, Praxair Gas). Preliminary experiments verified that growth rates of cultures bubbled at this rate were not significantly different from those of unbubbled cultures (data not shown), and these methods have been employed for other CO2 experiments (Hutchins et al. 2007; Fu et al. 2007), including dinoflagellate studies (Fu et al. 2008; 2010). Filtered seawater was amended with L1/20 nutrient, vitamin and trace metal concentrations (Guillard and Hargraves 1993), except NH4Cl+ was substituted for NaNO3- and silicate was omitted. Nutrient concentrations at the Redfield ratio (by atoms) of 16 N: 1 P (Redfield 1958) were added initially to the incubation bottles, and replenished once at the one week dilution (below).
The CO2-amended treatments were maintained in active growth using semi-continuous culture methods (Tatters et al. 2012). Each bottle was diluted to the original time-zero in vivo chlorophyll a fluorescence value after one week with nutrient-amended filtered seawater. Aliquots were removed initially, and after one and two weeks for examination of carbonate buffer system parameters and community structure using microscopic cell counts. Samples for cell counts were obtained at the 1 week time point (after dilution), and after the 2 week incubation, in order to calculate acclimated growth rates (1-2 week rates) and final abundances of all species.
Clonal culture isolations
Three individual cells representing the four dominant genera were isolated from each incubation bottle at the end of the two-week incubation of the natural community, and maintained in long-term culture (52 weeks) at the pCO2 from which they were obtained under conditions of temperature, light, nutrients, CO2 bubbling, etc., identical to the 2-week natural community experiment. Cultures were maintained in exponential phase using autoclave-sterilized enriched seawater growth medium with semi-continuous weekly dilutions based on specific growth rates within each bottle, calculated as in Tatters et al. (2012). The approximate number of generations during this time period was: L. polyedrum (48-62), P. micans (58-71), Alexandrium sp. (34-38) and Gonyaulax sp. (75-126).
Artificial community competition experiments
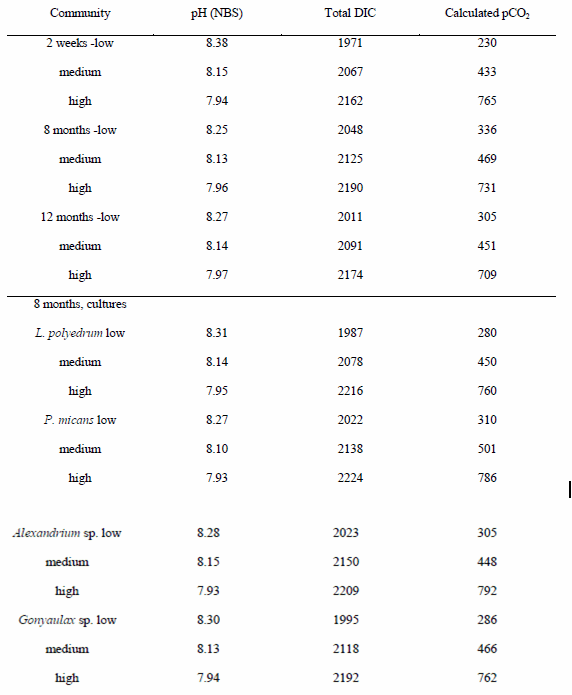
The conditioned clonal cultures were recombined into artificial communities after 4, 8 and 12 months in the same relative proportions and cell densities as the original natural bloom assemblage. The 8- and 12-month experiments used triplicate communities of all four species, but due to logistical limitations, the 4-month experiment used only L. polyedrum, P. micans, and Alexandrium sp. in duplicate communities. Because Gonyaulax sp. was not included in this preliminary 4-month experiment and replication was different, it is not fully comparable to the other experiments. The dinoflagellates in the artificial community trials were allowed to compete under identical experimental conditions of light, temperature, nutrient availability, and pCO2 for the same time period and diluted exactly as in the original natural bloom incubation. Samples were collected for cell counts and carbonate system parameters (Table 1) in all experiments.
The initial bloom counts, per mL, are as follows:
Lingulodinium: 464
Prorocentrum: 50.5
Alexandrium: 20.5
Gonyaulax: 27
Three samples (A, B, C) were taken for each partial pressure of CO2. The partial pressures of CO2 were targeted at 190, 390, and 750 ppm but the biology can interfere a bit with the actual values achieved.
Literature Cited
Fu, F.-X., M. E. Warner, Y. Zhang, Y. Feng, and D. A. Hutchins. 2007. Effects of increased temperature and CO2 on photosynthesis, growth and elemental ratios of marine Synechococcus and Prochlorococcus (Cyanobacteria). J. Phycol. 43:485-496.
Fu, F.-X., A. R. Place, N. S. Garcia, and D. A. Hutchins. 2010. CO2 and phosphate availability control the toxicity of the harmful bloom dinoflagellate Karlodinium veneficum. Aquat. Mcrob. Ecol. 59: 55-65.
Fu, F.-X., Y. Zhang, M. E. Warner, Y. Feng, and D. A. Hutchins. 2008. A comparison of future increased CO2 and temperature effects on sympatric Heterosigma akashiwo and Prorocentrum minimum. Harmful Algae. 7:76-90
Hutchins, D. A., F.-X. Fu, Y. Zhang, M. E. Warner, Y. Feng, K. Portune, P. W. Bernhardt, and M. R. Mulholland. 2007. CO2 control of Trichodesmium N2 fixation, photosynthesis, growth rates, and elemental ratios: Implications for past, present and future ocean biogeochemistry. Limnol. Oceanogr. 52:1293-1304.
Redfield, A. C. 1958. The biological control of chemical factors in the environment. Am. Sci. 46:205-222.
Tatters, A. O., F.-X. Fu, and D. A. Hutchins. 2012. High CO2 and silicate limitation synergistically increase the toxicity of Pseudonitzschia fraudulenta. PLoS ONE 7(2):e32116.doi:10.1371/journal.pone.0032116.
Analytical methods
Cell counts and growth rates. Growth rates were measured for each species in mixed communities during the second week of the initial natural community incubation, and during the 12-month and switch artificial community experiments. Because of logistical limitations and the fact that dinoflagellates cannot be cryopreserved for subsequent growth rate measurements, growth rates were determined immediately for all isolates in unialgal culture only at the 8-month timepoint. These values are representative of their in long-term steady-state exponential growth rates throughout the conditioning period. Final cell abundances of each species were measured in every natural or artificial community competition experiment. Algal cells were preserved in acidified Lugol’s solution and enumerated using an Accu-Scope 3032 inverted microscope using the Utermöhl method (Utermöhl, 1931).
Carbonate buffer system. Dissolved inorganic carbon analysis used a CM5230 CO2 coulometer (UIC) (King et al. 2011). pH was determined on freshly collected samples using a calibrated Orion 5-star plus pH meter using an NBS buffer system with three-point calibration. Experimental pCO2 was calculated using CO2SYS software as in Tatters et al. (2012) (Table 1). Due to unavoidable minor variability in calculated pCO2 levels between experiments (largely from differences in batches of commercial gas mixtures), for clarity, throughout the text pCO2 values of 230-336 ppm are referred to as “low”, values of 433-506 ppm are referred to as “medium”, and values of 709-792 ppm are referred to as “high”.
Multivariate statistical methods
Multivariate analyses used the PRIMER v6 statistics package (Clarke and Warwick, 2001) with the PERMANOVA add-on. Final cell abundances for each species from replicate bottles were square-root transformed prior to community structure comparisons based on Bray-Curtis similarities and log-transformed species growth rates compared based on Euclidean distance measures. ANOSIM permutation tests were used to test the impact of differing pCO2 competition levels on overall community structure and on relative abundance of the four species at the end of all incubations, and on their growth rates in the initial natural community, 12-month and switch experiments. These tests resulted in R values and significance levels where R = 0 implies no difference among groups, and R = 1 suggests that group separation is so large that all dissimilarities among groups are larger than any dissimilarity within them (Clarke and Warwick, 2001).
Data from the natural community experiment and from competition trials after 8 and 12 months were further combined for PERMANOVA analyses to test for significant differences among and within predefined groups in response to both the pCO2 competition levels and the differing periods of conditioning to these pCO2 concentrations. PERMANOVA resulted in Pseudo-F and significance levels, where Pseudo-F = 1 implies a large overlap among sample groups being compared while Pseudo-F > 1 indicates little or no overlap between them (Anderson et al., 2008). PERMANOVA also allowed us to test for interaction between the factors pCO2 competition level and conditioning period in forcing overall community structure.
For the 12-month switch competition trials, we used final cell abundances from all treatments to examine the comparative effects of differing pCO2 competition levels and differing pCO2 conditioning levels. Using a two-way crossed design for the ANOSIM routine, we tested the average effect on overall community structure and on the four individual species separately of pCO2 levels during competition removing differences in conditioning pCO2, and the average effect of conditioning pCO2 removing differences in competition pCO2 (Clarke and Warwick, 2001).
Literature Cited:
Anderson M. J., R. N. Gorley, and K. R. Clarke. 2008. PERMANOVA+ for PRIMER: Guide to software and statistical methods. PRIMER-E: Plymouth, UK.
Clark, K. R., and R. M. Warwick. 2001. Change in marine communities: an approach to statistical analysis and interpretation. 2nd edition. PRIMER-E, UK.
King, A.L., S. A. Sañudo-Wilhelmy, K. Leblanc, D. A. Hutchins, and F.-X. Fu. 2011. CO2 and vitamin B12 interactions determine bioactive trace metal requirements of a subarctic Pacific diatom. Intl. Society of Microb. Ecol. J.5(8):1388-1396.
Tatters, A. O., F.-X. Fu, and D. A. Hutchins. 2012. High CO2 and silicate limitation synergistically increase the toxicity of Pseudo-nitzschia fraudulenta. PLoS ONE 7(2):e32116.doi:10.1371/journal.pone.0032116.
Utermöhl, H.1931. Neue Wege in der quantitativen Erfassung des Planktons. (Mit besonderer Beriicksichtigung des Ultraplanktons). Verh. Int. Verein. Limnol. 5:567-596.
| File |
|---|
Dinoflagellate_Evo.csv (Comma Separated Values (.csv), 3.12 KB) MD5:fb637fe551c115a6160a1a698889e690 Primary data file for dataset ID 3796 |
| Parameter | Description | Units |
| incubation_period | Period of incubation | months |
| species | Species name | n/a |
| pCO2 | Partial pressure of carbon dioxide | |
| sample | Sample name, A, B, or C | n/a |
| growth_rate | Growth rate | divisions per day |
| count | Count of the cells | cell counts per mL |
| Website | |
| Platform | USC |
| Start Date | 2009-09-20 |
| End Date | 2010-12-31 |
| Description | Hutchins laboratory, University of Southern California
Comparing short vs. long term acclimation of a dinoflagellate community
Experiments conducted as part of project titled, "Experimental studies to understand and evaluate acclimation of marine plankton assemblages to increased CO2 and temperature". |
Progressing ocean acidification and increasing sea surface temperature may significantly impact marine plankton community structure and community-level processes. Yet, our ability to predict specific responses is still limited because of the tremendous taxonomic complexity of microbial assemblages and the limitations of the methodological and experimental tools presently available to test specific hypotheses. Research to study community level effects due to a changing CO2/temperature regime often involve short-term field incubations that subject organisms to simulated 'greenhouse' conditions. A central question for understanding global climate change is whether the trends and patterns that are observed in communities during short-term manipulations can be extrapolated to the responses of fully acclimated plankton communities over decadal or longer timescales.
The specific objectives of this research program are: 1) to examine how protistan communities restructure in response to increased seawater CO2 concentrations and temperature in semi-continuous field incubation experiments, and 2) to evaluate if the dominant algal species that are isolated from either ambient or increased CO2 and temperature treatments in field experiments will re-establish dominance under the same conditions in acclimated laboratory culture competition studies. Changes in community structure of natural protistan assemblages in our experimental treatments will be followed using image-based methods (flow cytometry, FlowCAM and microscopy) in combination with state-of the art molecular tools (DNA fingerprinting). Molecular approaches have begun to reveal an incredible high diversity for marine microbes and stimulate debate in regard to the ubiquitous presence of a microbial 'Rare Biosphere' that is, the presence of a huge number of species that are present at extremely small percentages of the total abundance of microbes, among a much smaller percentage of dominant ones. Little is known about the ecological significance of these rare species, and the investigators hypothesize that change in CO2 and temperature will select for some of these members that are inconspicuous under ambient conditions.
The unique aspect of this experimental approach is the combined use of field incubations that encompass entire natural microbial assemblages, with a series of laboratory culture competition trials that focus on the same groups of algae after extended acclimation, to evaluate the validity of short-term experiments that examine changing CO2 and temperature. First, field incubation experiments will be conducted to characterize changes in protistan community structure under ambient and future CO2/temperature regimes. Second, clonal algal strains will be isolated from dominant taxa in present day and greenhouse treatments, and cultivated for extended periods under their 'preferred' CO2/temperature conditions. Finally, mixtures of these acclimated strains will be competed against each other, to re-examine their responses to ambient and greenhouse conditions and compare them to the responses observed in the unacclimated field incubation experiments.
Two graduate students will make this project the focus of their Ph.D. research at USC, and undergraduate students will be involved in the field and laboratory work. Results from this research will be incorporated in lesson plans on microbial diversity and global climate change. Dissemination of data and results is planned on a project website. The PIs in this project also participate in an on-going, innovative, NSF-funded program (Centers for Ocean Science Education Excellence; COSEE-West) which focuses on personal involvement of faculty in a custom framework to allow an effective connection with K-12 teachers, thus improving math and science education in disadvantaged parts of Southern California.
| Funding Source | Award |
|---|---|
| NSF Division of Ocean Sciences (NSF OCE) |
{kind=link}